
神经元(neuron)是神经系统中的基本结构单位。一个典型的神经元由胞体和突起两部分组成(见图)。胞体又包括胞核和核周围胞质两部分。神经元的胞核很大,从容积或表面积看,它占胞体的大部分。而核周体虽然只占小部分,但它对维持整个神经元的代谢和功能起着重要的作用。胞体是合成各种蛋白质(包括各种酶类)的中心。蛋白质首先在胞体中合成,然后再经过运输系统运送到突起之中。
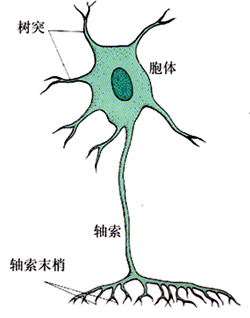
突起可以分为树突和轴突。树突的分支较短,由胞体发出后逐渐变细,并不断分支。树突可以看成是胞体的延伸部分。轴突一般较长,由胞体发出后一般不分支,直到轴突的末端才形成终末侧支。也有少数神经元的轴突在到达终末侧支前分出侧索。轴突一般都有髓鞘(myelin)包被。在轴突与胞体连接部以及轴突的终末侧支失去髓鞘。前者称为轴突的起始段,后者称为纤细终末。起始段的兴奋性最高,往往是形成冲动的部位。轴突内的细胞质称为轴浆(axoplasm),内有神经细丝、微管、线粒体、囊泡等,但不能合成蛋白质。因此,一些合成神经递质的酶或肽神经递质,往往是在胞体合成后经轴浆运输到达轴突的末梢。
由于神经元的轴突很长,又称为神经纤维。根据神经纤维的直径大小及来源,可将其分成Ⅰ、Ⅱ、Ⅲ、Ⅳ类。也可根据神经纤维上冲动的传导速度及电生理特性分成A、B、C三大类。神经纤维上有的有髓鞘,有的无髓鞘(实际上有一层很薄的髓鞘),因此又可以分成有髓鞘和无髓鞘两类。还可以从神经纤维传导冲动的方向将其分成传入神经和传出神经两类。这些分类之间的关系,往往使初学者容易造成混乱。表1和表2总结了不同分类法神经纤维的特点和它们之间的关系。
| 表1 神经纤维的分类(一) | ||||||||
| 纤维分类 | A 类(有髓纤维) | B类 (有髓纤维) |
C类(无髓纤维) | |||||
| Aα | Aβ | Aγ | Aδ | sC | drC | |||
| 来源 | 初级肌梭传入纤维和支配梭外肌的传出纤维 | 皮肤的触压觉传入纤维 | 支配梭内肌的传出纤维 | 皮肤痛温觉传入纤维 | 自主神经节前纤维 | 自主神经节后纤维 | 后根中传导痛觉的传入纤维 | |
| 纤维直径(μm) | 13~22 | 8~13 | 4~8 | 1~4 | 1~3 | 0.3~1.3 | 0.4~1.2 | |
| 传导速度(m/s) | 70~120 | 30~70 | 15~30 | 12~30 | 3~15 | 0.7~2.3 | 0.6~2.0 | |
| 锋电位持续时间(ms) | 0.4~0.5 | 1.2 | 2.0 | |||||
| 负 后 电 位 |
% 锋电位高度 | 3~5 | 无 | 3~5 | 无 | |||
| 持续时间(ms) | 12~20 | - | 50~80 | - | ||||
| 正 后 电 位 |
% 锋电位高度 | 0.2 | 1.5~4.0 | 1.5 | 10~30 | |||
| 持续时间(ms) | 40~60 | 100~300 | 300~400 | 75~100 | ||||
| 表2 神经纤维的分类(二) | ||||
| 纤维类别 | 来源 | 直径(μm) | 传导速度(m/s) | 电生理学上的分类 |
| Ⅰ | 肌梭及腱器官的传入纤维 | 12~22 | 70~120 | Aα |
| Ⅱ | 皮肤的机械感受器传入纤维(触、压、振动感受器传入纤维) | 5~12 | 25~70 | Aβ |
| Ⅲ | 皮肤痛温觉传入纤维,肌内的深部压觉传入纤维 | 2~5 | 10~25 | Aδ |
| Ⅳ | 无髓的痛觉纤维,温度、机械感受器传入纤维 | 0.1~1.3 | 1 | C |
冲动在神经纤维上的传导,是一种局部电流的传导(图)。神经纤维兴奋部位,由于在动作电位形成时,Na+由膜外快速进入膜内,造成膜的外表面的电位降低。而邻近神经纤维膜的外表面的电位,由于仍为安静时的静息电位,其电位较高,膜的表面又都附有组织液,可以导电,因此,兴奋部位与安静部位之间形成一种局部电流。其电流方向,在细胞膜的外表面是由安静部位流向兴奋部位;在神经纤维内,其方向相反。在膜内,这个电流可提高膜内电位。当安静部位电流的强度不断上升,达到阈电位时,可打开膜的电压门控Na+通道,使安静部位产生动作电位。兴奋则由一处传到了另一处。这种传导的方式在神经纤维上具有以下几个特点。
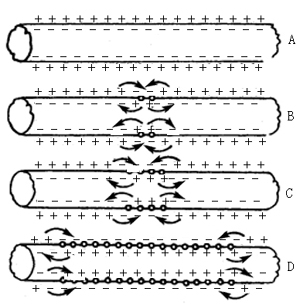
1.结构和功能完整性 冲动的传导必须是在一根完整的神经纤维上才能进行。如果神经纤维被折断,局部电流则很难由断端部位向前传导。而如果神经纤维的一部分由于药物或毒素使其丧失功能,不能产生动作电位,尽管在形态上是完整的,也不能传导冲动。因此,神经纤维在解剖和生理上的完整性是冲动传导的必需条件。
2.绝缘性 一条神经干中有很多的神经纤维。例如人的坐骨神经中有多达20多万根神经纤维。每条神经纤维的动作电位(或冲动)的传导是互相不干扰的,这是因为神经纤维上都有一层髓鞘,起着绝缘作用。
3.传导速度 冲动的传导实际上是局部电流的传导,它的传导速度与神经纤维的粗细、髓鞘的厚薄、局部电流强度等有关。神经纤维越粗,传导速度越快,局部电流强度越大传导速度越快。最快的A类纤维,其传导速度可超过100m/s。尽管神经传导速度可以很快,但与电流传导速度(3×106m/s)相比,还是很慢的。
在有髓鞘的神经纤维中,实际上往往髓鞘并不是将神经纤维全部包裹起来,而有结——郎飞氏结。在结处,髓鞘很薄,电阻最小。当形成局部电流时,结处的电流强度最大,最易引起去极化产生动作电位。因此,局部电流的传导,是由一个结传给另一结作跳跃式的传导,称为跳跃传导。这种传导方式可以加快冲动的传导速度。这也是有髓鞘或有郎飞氏结的纤维具有较快传导速度的原因。
4.双向传导 将一根神经纤维分离出来,在神经纤维的中间给一个电刺激,产生兴奋,形成动作电位。这一动作电位的形成,使膜的外表面电位较周围静息部位低(由Na+快速进入膜内造成)。 因此,兴奋部位两侧的静息部位都可以与兴奋部位产生局部电流。只要电流强度能使安静部位膜电位的变化达到阈电位,都能打开快速钠通道(Na+通道)而形成动作电位,产生兴奋。因此,形成的冲动可由刺激部位向相反的两个方向传导,即冲动传导的双向传导。在整体条件下,由于冲动往往都是由树突或胞体向轴突方向传导,而在化学性突触部位,其传导方向是单向而非双向。因此,在整体条件下一般不存在双向传导。只是在某些特别情况下(如轴突反射)有双向传导的可能性。
5.相对不疲劳性 冲动的传导实际上是通过局部电流进行的传导。局部电流形成的条件是兴奋部位神经纤维表面电位降低,与周围静息部位形成电位差产生电流。这完全是一种物理现象,无需直接提供能量。兴奋部位电位降低的原因,是由于钠通道打开,Na+由膜外快速流入膜内。Na+的流动是由高浓度的膜外流向浓度低的膜内,这一运动方式也是一种无需直接供能的活动。因此冲动在神经纤维上的传导本身基本上是一种无需供能的活动,与突触部位的传导相比,不易产生疲劳。
6.不衰减性 冲动传导的机理是局部电流的传导,而不是电流本身的传导。兴奋部位电位的降低取决于膜上钠通道的打开,形成锋电位。只要形成锋电位的条件不变,锋电位的大小形状是不会改变的。因此不会因为神经纤维很长电阻加大而降低锋电位,即传导的不衰减性。
神经元的胞体是合成蛋白质(包括酶)的部位。合成后的蛋白质可以通过转运系统输送到轴突的末端。例如以释放肽类物质为其神经递质的神经元,这些肽类神经递质的前体首先在胞体合成后再转运到轴突的远端。一些释放经典神经递质的神经元,这些神经递质如5-HT等,虽然可以在神经末梢合成,但合成这些神经递质的酶不能在神经末梢中合成,必须由胞体合成后再转运到末梢。神经纤维中构成微管、微丝等细胞骨架的蛋白质以及一些与之有关的蛋白质,也需要首先在胞体合成后再运输到神经纤维的末梢。
神经末梢还可吸收一些蛋白质,通过反方向的转运送到胞体。因此,在正常条件下,神经纤维中存在着两种方向相反的运输过程,顺向转运和逆向转运。
轴浆运输的速度可快可慢,不同的物质具有不同的速度。膜蛋白、分泌蛋白、膜磷脂以及胆固醇等的转运速度较快。目前认为这类快速转运的物质不是以单分子形式转运,而是经过包装后以囊泡或囊泡小管形式转运。快速转运的速度可达300~400mm/d。一些与细胞骨架有关的微管、微丝蛋白以及一些酶类的转运速度则很慢,速度只有0.01~1mm/d。
轴浆运输是需要代谢提供能量的,例如在缺氧条件下快速转运减慢甚至中止。轴浆运输也是可以调节的,例如在神经被切断后,一些与生长有关的蛋白质的转运可增加100倍。关于轴浆 运输的详细机理目前还了解不多。
逆向运输,即由末梢向胞体的转运。这类转运的物质包括一些可供神经元再利用的物质,经末梢吸收后转运至胞体。破伤风毒素及狂犬病毒由外周进入中枢也可能是利用了这种逆向运输机制。辣根过氧化酶可以由神经末梢吸收后通过逆向运输机制向胞体运输。这种酶可促进某些底物的氧化而着色,因此在一部位注射辣根过氧化酶后,在它的传送过程中以及最后到达胞体,都可以作连续切片,从形态上追踪该酶的去向。这一方法已被广泛应用于神经系统的研究。